
Sản xuất nông nghiệp là hoạt động của con người không chỉ đảm bảo lượng lương thực, thực phẩm cần thiết cho nhu cầu cuộc sống, mà còn làm tăng đáng kể lượng hoá chất gây ô nhiễm vào môi trường thông qua việc sử dụng quá nhiều phân bón hoá học, thuốc trừ sâu, diệt cỏ. Những chất này không chỉ gây hại cho môi trường, mà chúng còn là các nguy cơ tiềm ẩn đối với sức khoẻ của con người, vật nuôi và môi trường đất, nước, không khí [1]. Người nông dân bón quá nhiều phân bón nitrate ammon (NH4NO3) với mục đích tăng năng suất cây trồng, tuy nhiên việc làm này dẫn đến sự suy giảm khả năng cố định nitơ sinh học của vi sinh vật trong đất vì lượng NH+4 trong đất dư thừa, cây trồng không cần các vi sinh vật cố định nitơ sinh học để cung cấp ammon nữa. Lượng ammon dư thừa được các vi sinh vật nitrate hoá chuyển thành N2O bởi các vi sinh vật phản nitrate hoá hoặc ngấm vào nước ngầm gây nên hiệu ứng nhà kính hoặc ô nhiễm nước ngầm. Chính vì thế, sản xuất nông nghiệp cần phải tìm được giải pháp thích hợp để giải quyết các vấn đề do sử dụng phân bón, thuốc trừ sâu, bảo vệ thực vật hoá học và giảm sự phụ thuộc vào các chất này. Một trong những biện pháp đó là khai thác, sử dụng các PGPR. PGPR là thành phần quan trọng đóng góp vào độ phì nhiêu của đất, kích thích cây trồng tăng trưởng và có hiệu quả đối kháng với các tác nhân gây bệnh hại cây trồng [2] và không gây ô nhiễm môi trường [3] .
Vùng rễ (rhizosphere) là vùng đất xung quanh bề mặt rễ cây – nơi các hoạt động sinh học của vi sinh vật diễn ra mạnh nhất [4]. PGPR là các vi sinh vật sinh trưởng và phát triển trong đất, xâm chiếm bề mặt rễ cây và có hiệu quả kích thích cây trồng sinh trưởng [5]. PGPR tham gia vào các hoạt động sinh học trong đất như phân huỷ các tàn dư thực vật, tổng hợp các chất hữu cơ, khoáng hoá chất hữu cơ, cố định các chất dinh dưỡng, do đó góp phần tăng cường và cải thiện độ phì nhiêu của đất. Nhiều vi khuẩn, vi nấm, xạ khuẩn có vai trò kích thích sinh trưởng cây trồng đã được thương mại hoá như Pseudomonas, Azospirillum, Azotobacter, Bacillus Xanthomonas…, tuy nhiên việc khai thác và sử dụng có hiệu quả PGPR vẫn chỉ chiếm một phần rất nhỏ trong sản xuất nông nghiệp. Vấn đề này có thể do các đặc tính của PGPR khi được sử dụng trên đồng ruộng biểu hiện không đồng đều, bền vững nên có thể ảnh hưởng tới sinh trưởng của cây trồng. Sử dụng hiệu quả các chế phẩm sinh học từ PGPR phụ thuộc trước hết vào sự tồn tại của chủng PGPR trong đất, tiếp đến là khả năng tương tác với cây trồng, các vi sinh vật bản địa trong đất, các yếu tố môi trường. Mặt khác, do cơ chế tác động của PGPR rất đa dạng và không phải tất cả PGPR đều có chung cơ chế tác động, nên PGPR cần phải có khả năng cạnh tranh cao tại vùng rễ, thúc đẩy tăng trưởng thực vật thông qua nhiều cơ chế tác động, tương thích với các chủng vi sinh vật khác tại vùng rễ, chịu được tác động của nhiệt độ cao, an toàn với người sản xuất và môi trường. Theo quan điểm sản xuất nông nghiệp bền vững, trong tương lai, các chế phẩm sinh học từ PGPR sẽ được sử dụng rộng rãi trong sản xuất.
Cơ chế tác động của PGPR
PGPR có khả năng tăng cường sinh trưởng của cây trồng thông qua cơ chế tác động trực tiếp hoặc gián tiếp. Cơ chế trực tiếp gồm hoà tan phosphate, cố định nitơ, tổng hợp siderophore, HCN, phytohormones (như auxin, cytokinin và gibberellin), ngược lại, cơ chế gián tiếp gồm hoạt hoá ACC deaminase, tổng hợp antibiotics và các enzyme thuỷ phân, tăng cường tính kháng hệ thống [1, 5].
Cố định nitơ
Nitơ là chất dinh dưỡng quan trọng với sinh trưởng và năng suất của cây. Khoảng 78-80% nitơ có trong không khí, nhưng cây trồng không sử dụng được nguồn nitơ này. Nitơ trong không khí được chuyển thành dạng dễ hấp thụ với cây trồng thông qua quá trình cố định nitơ bởi các vi sinh vật sử dụng enzyme nitrogenase. Các vi sinh vật cố định nitơ thường được phân loại thành: vi sinh vật cộng sinh cố định nitơ (gồm các vi khuẩn cộng sinh với cây họ đậu); các vi sinh vật sống tự do như Azospirillum, Azotobacter, Gluconoacetobacter diazotrophicus. Lượng nitơ được cố định bởi các vi sinh vật tự do rất nhỏ so với lượng nitơ được cố định bởi vi sinh vật cộng sinh.
Hoà tan phosphate
Phospho là một trong các nguyên tố cần thiết với sinh trưởng và phát triển của cây trồng. Phospho tham gia nhiều vào quá trình trao đổi chất quan trọng của cây như quang hợp, hô hấp, chuyển năng lượng [6]. Lượng lớn phopspho tồn tại ở dạng hữu cơ và vô cơ không tan trong đất. Do đó, vi sinh vật hoà tan phosphate (Phosphate solubilizing bacteria – PSB) đóng vai trò quan trọng trong quá trình giải phóng phosphate từ các phân tử hữu cơ hoặc hoà tan các hợp chất phosphate vô cơ khó tan. Thực vật hấp thụ phosphate ở dạng ion H2PO4– hoặc H2PO42-Cơ chế hoà tan phosphate bởi các PSB liên quan tới giải phóng các acids hữu cơ khối lượng phân tử thấp như acetic, lactic, oxalic…, thông qua các nhóm hydroxyl và carboxyl chúng kéo các cations bao quanh nhóm phosphate, cuối cùng chuyển hợp chất phosphate khó tan thành dạng tan, dễ hấp thụ bởi cây trồng. Các vi sinh vật có khả năng phân giải các hợp chất phosphate khó tan trong đất được xếp vào nhóm các vi sinh vật phân giải phosphate (Phosphate solubilizing microorganisms – PSM).
Trong số các PSMs cư trú tại vùng rễ, PSB được coi là nguồn phân bón tiềm năng vì chúng có thể cung cấp cho cây trồng nguồn phospho dễ tiêu từ các nguồn phosphate khó tan trong đất bằng nhiều cách. Các chủng vi khuẩn thuộc các chi như Azotobacter, Bacillus, Beijerinckia, Burkholderia, Enterobacter, Erwinia, Flavobacterium, Microbacterium, Pseudomonas, Rhizobium và Serratia đã được báo cáo là các chủng PMS khó tan. Quá trình hoà tan cá hợp chất phospho vô cơ diễn ra nhờ tác động của các acid hữu cơ có khối lượng phân tử thấp, chúng được tổng hợp bởi nhiều vi sinh vật đất. Khoáng hoá các hợp chất phospho hữu cơ diễn ra thông qua tổng hợp các enzyme phosphatase khác nhau xúc tác phản ứng thuỷ phân cầu nối phospho-ester. Bên cạnh việc cung cấp phospho cho cây, PSB còn kích thích hoạt động của quá trình cố định nitơ, tổng hợp các cơ chất kích thích cây trồng tăng trưởng.
Tổng hợp siderophore
Sắt là nguyên tố cần thiết với nhiều hoạt động sống của tế bào. Tất cả vi sinh vật đã được biết đều cần sắt. Trong điều kiện hiếu khí, sắt tồn tại chủ yếu ở dạng Fe3+ và dễ tạo dạng hydroxide không tan, do đó không thích hợp cho thực vật và vi sinh vật. Vi khuẩn thu nhận sắt bằng cách tiết ra các hợp chất chelators sắt được gọi là siderophore có ái lực cao với sắt. Một số siderophore hoà tan trong nước và có thể được phân loại thành siderophore ngoại bào và nội bào. Nhóm rhizobacteria gram âm và gram dương, sắt Fe3+ tạo phức với siderophore trên màng tế bào vi khuẩn và được khử thành Fe2+ và giải phóng vào trong tế bào thông qua kênh nối giữa màng ngoài và trong của tế bào. Trong quá trình này, siderophore có thể được tái sử dụng hay phá hủy. Siderophore đóng vai trò như tác nhân giải phóng sắt khỏi các thành phần khoáng và hữu cơ trong điều kiện thiếu sắt. Nhiều nhà nghiên cứu đã thông báo các chủng vi khuẩn tổng hợp siderophore có vai trò kích thích sinh trưởng thực vật và kiểm soát sinh học.
Tổng hợp các phytohormone
Khoảng 80% vi sinh vật vùng rễ của nhiều loại cây trồng khác nhau có khả năng tổng hợp và giải phóng các hợp chất auxin như là sản phẩm trao đổi chất thứ cấp. Indol-3-acetic acid (IAA) được tiết ra bởi vi sinh vật vùng rễ tác động vào nhiều quá trình phát triển của thực vật do nguồn dự trữ IAA trong thực vật có thể thay đổi do thu nhận IAA được tiết ra bởi vi khuẩn trong đất. IAA từ vi khuẩn làm tăng diện tích bề mặt và chiều dài rễ, do đó giúp thực vật thu nhận được nhiều dinh dưỡng trong đất. IAA từ vi sinh vật vùng rễ làm mềm thành tế bào thực vật nên tăng lượng chất tiết ra từ rễ, bổ sung thêm nguồn dinh dưỡng cho vi sinh vật tại vùng rễ. Hợp chất làm thay đổi nồng độ IAA là tryptophan – tiền chất cần thiết của quá trình tổng hợp IAA. Tryptophan tăng tổng hợp IAA, ngược lại anthranilate – tiền chất tổng hợp tryptophan lại giảm tổng hợp IAA. Acinetobacter, Pseudomonas, Rhizobium, Azospirillum, Bacillus và Klebsiella là các chi vi khuẩn thường tham gia vào tổng hợp IAA.
Gibberellin rất quan trọng với sự nảy mầm của hạt, cảm ứng ra hoa, phát triển của hoa và quả, hiệu quả sinh lý trội nhất của gebberellin là kéo dài chồi. Gebberellin được tổng hợp bởi thực vật bậc cao, vi nấm và vi khuẩn. Nhiều chi vi khuẩn tổng hợp gibberellin được công bố như Acetobacter diazotrophicus, Azospirillum lipoferum, Bacillus pumilus, Bacillus cereus, Bacillus macrolides và Herbaspirillum seropedicae [6].
Ethylene là phytohormone có chức năng sinh học quan trọng đối với sinh trưởng và phát triển của thực vật. Nó kích thích sự ra rễ, ức chế kéo dài rễ, giảm sự héo rũ, thúc đẩy sự chín của quả, thúc đẩy sự nảy mầm của hạt, kích thích tổng hợp các hormone thực vật khác. Sự có mặt của enzyme 1-aminocyclopropane-1-carboxylate (ACC) deaminase được tổng hợp bởi PGPR góp phần điều hoà nồng độ ethylene trong thực vật. Các nhóm vi khuẩn thuộc chi Pseudomonas, Bacillus, Acinetobacter, Azospirillum, Achromobacter, Enterobacter, Agrobacterium, Rhizobium… đã được chứng minh có khả năng tổng hợp ACC – deaminase.
Tổng hợp HCN và chất kháng sinh
HCN thường được sử dụng như tác nhân kiểm soát sinh học trong sản xuất nông nghiệp dựa trên độc tính kháng lại các tác nhân gây bệnh hại thực vật, chelate các ion kim loại và tham gia gián tiếp vào quá trình tạo ra các hợp chất phosphate dễ hấp thụ bởi thực vật. Aeromonas, Pseudomonas, Bacillus và Enterobacte là các chi vi khuẩn liên quan tới tổng hợp HCN. Nhiều chủng vi khuẩn PGPR tổng hợp chất kháng sinh (antibiotics) ức chế sự phát triển của các mầm bệnh. Các loài vi khuẩn PGPR như Bacillus subtilis, Bacillus amyloliquefaciens, Bacillus velezensi, P.putida, P.fluorescens, Pseudomonas brassicacearum và Paenibacillus polymyxa tổng hợp nhiều antibiotics khác nhau như surfactin, bacillomycin, fengycin, iturin, 2,4-diacetylphloroglucinol,polymyxin và fusaricidin là chất kháng sinh ức chế mạnh sự phát triển của Ralstonia solanacearum [6].
Tăng cường tính kháng hệ thống
PGPR kích thích tính kháng hệ thống tiềm ẩn của thực vật, do đó cải thiện tính kháng của cây chống lại các mầm bệnh. Ví dụ, Bacillus amyloliquefaciens, actobacillus paracasei, P. fluorescens và P. putida cảm ứng tính kháng hệ thống của cây chống lại các mầm bệnh hại cây.
Các enzyme thuỷ phân
Các enzyme thủy phân như urease, esterase, lipase,protease, chitinase, amylase và cellulose rất quan trọng trong quá trình chuyển hoá các hợp chất carbon, nitơ và hydro. Các enzyme thuỷ phân tham gia phá huỷ thành tế bào của vi nấm, làm thay đổi cấu trúc, thành phần thế của chúng. Đây chính là một cơ chế thân thiện với môi trường để phòng trừ nấm gây bệnh hại cây. Nhiều loài vi khuẩn như Serratia marcescens, Bacillus cereus, Bacillus thuringiensis tổng hợp các enzyme thủy phân và được ứng dụng trong kiểm soát sinh học các mầm bệnh như Rhizoctonia solani, Fusarium oxysporum, Sclerotium rolfsii, Pythium ultimum bằng cách làm phồng, sưng sợi nấm và sưng ở đầu sợi nấm, bẻ cong sợi nấm hay phá vỡ đầu sợi nấm của nấm gây bệnh.
PGPR tác động như phân bón sinh học
Phân bón sinh học là chế phẩm được sản xuất từ các chủng vi sinh vật hữu ích đã được tuyển chọn, đánh giá và hỗn hợp các cơ chất có khả năng phân huỷ sinh học. Các chủng vi sinh vật này có thể được sử dụng để tăng cường và cải thiện độ phì nhiêu của đất, năng suất của cây trồng. Khi được sử dụng cùng với hạt để bón vào đất, các chủng vi sinh vật sẽ xâm chiếm vùng rễ, biểu hiện các đặc tính có lợi và tăng cường sinh trưởng của cây, do đó nâng cao năng suất cây trồng. Các tương tác có lợi giữa cây trồng – vi sinh vật là giải pháp có tính bền vững để cải thiện năng suất cây trồng và thay thế dần phân bón và thuốc bảo vệ thực vật nguồn gốc hoá học [7]. Phân bón sinh học và thuốc trừ sâu sinh học là các sản phẩm có nguồn gốc tự nhiên được sử dụng rộng rãi để nâng cao chất lượng đất và được sử dụng như tác nhân kiểm soát sinh học. PGPR như phân bón sinh học cung cấp giải pháp an toàn, hiệu quả và thân thiện với môi trường để tăng năng suất cây trồng, chất lượng sản phẩm. Hiện nay, đã có nhiều chủng vi sinh vật thuộc các chi khác nhau được phân lập từ vùng rễ cây hoặc từ trong cây (vi khuẩn nội cộng sinh) được sử dụng như phân bón sinh học và chất trừ sâu sinh học. Các chủng vi khuẩn thuộc các chi như Azotobacter, Bacillus, Klebsiella, Enterobacter, Arthrobacter, Burkholderia, Bacillus spp. được khai thác sử dụng như phân bón sinh học đã được nhiều nhà khoa học công bố.
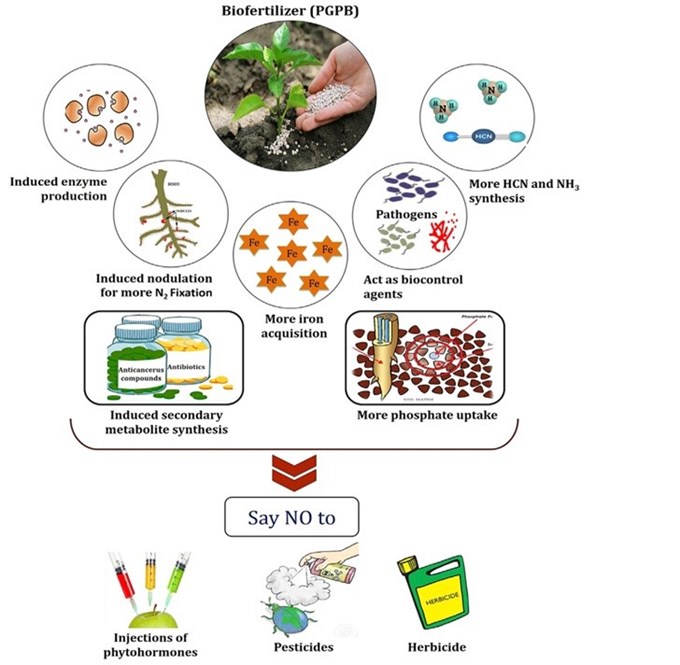 |
| Tác động của phân bón sinh học tới cây trồng và ức chế mầm bệnh [8]. |
KẾT LUẬN
Mặc dù thuốc trừ sâu và phân bón hoá học vẫn đang được sử dụng nhiều trong sản xuất nông nghiệp do dễ sử dụng, hiệu quả nhanh, nhưng chúng cũng là mối nguy tiềm ẩn với môi trường cũng như của chính sức khoẻ của con người. Vi sinh vật kích thích sinh trưởng thực vật/PGPR có nhiều hoạt tính có lợi cho cây trồng. Sử dụng PGPR như phân bón sinh học, thuốc trừ sâu sinh học được xem là biện pháp bền vững trong sản xuất nông nghiệp do phân bón sinh học ít hoặc không ảnh hưởng tới môi trường, và nâng cao năng xuất cây trồng.
PGPR giúp cây trồng kháng lại một số stress môi trường như hạn, mặn. PGPR được kỳ vọng sẽ thay thế phân bón và thuốc trừ sâu hoá học. Hiệu quả của PGPR có thể được tăng cường thông qua tối ưu hoá quá trình sản xuất, bảo quản và thích nghi với nhiều điều kiện thổ nhưỡng khác nhau. Chính vì vậy, có nhiều nghiên cứu về cơ chế tác động của PGPR để tuyển chọn được nhiều chủng vi sinh vật có thể hoạt động được trong nhiều điều kiện sinh thái, sản xuất nông nghiệp khác nhau.
TÀI LIỆU THAM KHẢO
[1] B.N. Aloo, et al. (2019), “The potential of bacilli rhizobacteria
for sustainable crop production and environmental sustainability”, Microbiol Res., 219, pp.26-39.
[2] A. Méndez-Bravo, et al. (2018), “Plant growth-promoting rhizobacteria associated with avocado display antagonistic activity against Phytophthora cinnamomi through volatile emissions”, PLOS ONE, 13(3), pp.1-13.
[3] P. Calvo, et al. (2014), “Agricultural uses of plant biostimulants”, Plant Soil, 383, pp.3-41.[4] M. Choudhary, et al. (2018), Towards Plant-Beneficiary Rhizobacteria and Agricultural Sustainability (Role of Rhizospheric Microbes in Soil), Springer, pp.1-46.
[5] H.P. Parewa, et al. (2018), Sustainable Crop Production
and Soil Health Management Through Plant Growth-Promoting Rhizobacteria (Role of Rhizospheric Microbes in Soil), Springer, pp.299-329.
[6] H. Mekonnen, M. Kibret (2021), “The roles of plant growth promoting rhizobacteria in sustainable vegetable production in Ethiopia”, Chem. Biol. Technol. Agric., 8, pp.1-11.
[7] S. Timmusk, et al. (2017), “Perspectives and challenges of microbial application for crop improvement”, Front Plant Sci., 8, DOI: 10.3389/fpls.2017.00049.
[8] Monika Singh, et al. (2019), Chapter three – Plant Growth Promoting Rhizobacteria: Application in Biofertilizers and Biocontrol of Phytopathogens (PGPR Amelioration in Sustainable Agriculture, Woodhead Publishing, pp.41-66.