
1. ĐẶT VẤN ĐỀ
Nấm linh chi (Ganoderma lucidum (Leyss. ex Fr.) Karst) là một loại thảo dược quý trong y học cổ truyền và được sử dụng làm thực phẩm chức năng nhằm phòng chống một số bệnh hiểm nghèo, các bệnh liên quan đến hệ tuần hoàn, hệ tiêu hóa [9].
Hiện nay, có nhiều phương pháp được sử dụng để chọn tạo giống nấm gồm: đột biến, dung hợp tế bào trần, chuyển gen và lai [7]. Trong đó, đột biến bằng tia gamma được coi là phương pháp hữu dụng để tạo ra các chủng đột biến cho năng suất cao [4]. Ứng dụng phương pháp đột biến bằng tìa gamma trong chọn tạo giống nấm đã thành công trên nhiều đối tượng. Chủng nấm sò (P. sajor-caju, P. columbinus và P. hybrid) đột biến bằng tia gamma và tia UV có thể cho năng suất cao và sinh trưởng tốt trong khoảng nhiệt độ rộng [7]. Đột biến bằng tia gamma có thể tạo ra các chủng nấm sò Pleurotus ostreatus có mức độ tương đồng di truyền với chủng đối chứng từ 64 đến 93%. Một số chủng đột biến có hoạt tính enzyme ngoại bào phân giải cellulose cao hơn 10 lần so với chủng đối chứng ban đầu [3]. Chủng nấm sò P. floridae đột biến bằng tia gamma 60Co có tốc độ sinh trưởng hệ sợi và năng suất sinh học cao hơn [2]. Chủng nấm mỡ (A. bisporus) đột biến tăng hàm lượng phenol tổng số so với chủng đối chứng sau 9 ngày bảo quản [1].
Trong nhiều năm qua, cùng với sự phát triển của khoa học kỹ thuật, công nghệ nhân giống và nuôi trồng nấm linh chi ở nước ta đã có những bước tiến nhất định. Tuy nhiên, do quá trình khai thác giống nấm linh chi trong thời gian dài đã làm ảnh hưởng đến sức sinh trưởng, khả năng chống chịu và có hiện tượng thoái hóa giống. Cho đến nay, công tác chọn tạo giống để thu được các chủng linh chi với nhiều tính trạng ưu việt như năng suất cao, giá trị dược liệu tốt và có khả năng thích nghi nhiều vùng sinh thái vẫn còn nhiều hạn chế và chưa đạt được những kết quả đáng kể. Với mục tiêu chọn tạo các chủng nấm linh chi đột biến tiềm năng cho năng suất tốt và giá trị dược liệu cao, 8 chủng đột biến bằng tia gamma được nghiên cứu về đặc điểm sinh trưởng hệ sợi, hình thành quả thể và năng suất.
2. KẾT QUẢ VÀ THẢO LUẬN
2.1. Đặc điểm hình thái và khả năng sinh trưởng hệ sợi của các chủng linh chi đột biến trên môi trường PGA
Để đánh giá khả năng sinh trưởng hệ sợi, các chủng linh chi đột biến và giống đối chứng được nhân trên môi trường PGA. Đường kính và tốc độ sinh trưởng hệ sợi được thể hiện ở Hình 1.
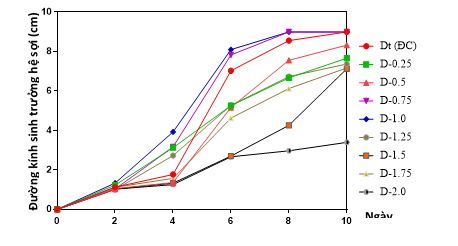 |
Hình 1. Sinh trưởng hệ sợi của các chủng linh chi trên môi trường cấp I
Dựa vào kết quả thu được có thể thấy cường độ chiếu xạ ảnh hưởng đến khả năng sinh trưởng hệ sợi của linh chi. Hai chủng đột biến D-0,75 và D-1,0 có khả năng sinh trưởng, phát triển hệ sợi tốt hơn so với công thức đối chứng (Dt). Tại ngày nuôi sợi thứ 6, đường kính hệ sợi của chủng đột biến D-0,75 và D-1,0 lần lượt là 7,38 cm và 8,1 cm, cao hơn giống đối chứng Dt (7,03 cm) (hình 1 và hình 2). Tại ngày nuôi sợi thứ 8, đường kính hệ sợi của hai chủng đột biến sinh trưởng kín bề mặt môi trường nhân giống, đường kính hệ sợi của hai chủng cùng đạt 9 cm (giống đối chứng Dt đạt 8,56 cm). Mật độ hệ sợi của hai chủng đột biến D-0,75 và D-1,0 được đánh giá dày, phân bố đều trên môi trường nuôi cấy thuần nhất (hình 3). Các chủng D-1,5, D-1,75 và D-2,0 với cường độ chiếu xạ cao có tốc tộc sinh trưởng hệ sợi chậm, phát triển không đồng đều trên môi trường; hệ sợi nhanh hình thành mô sẹo.
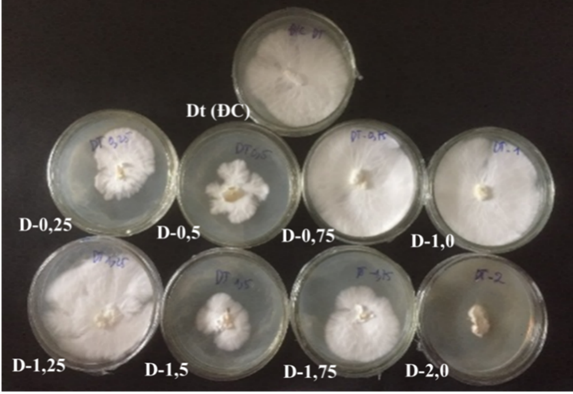 |
Hình 2. Sự phát triển hệ sợi của chủng nấm linh đột biến sau 6 ngày nuôi ở nhiệt độ 28oC
2.2. Khả năng sinh trưởng, phát triển của các chủng linh chi đột biến trên môi trường nhân giống cấp II
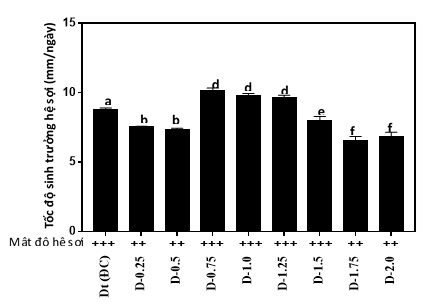 |
Hình 3. Tốc độ sinh trưởng và mật độ hệ sợi các chủng linh đột biên trên môi trường nhân giống cấp II
Kết quả nghiên cứu đặc điểm sinh trưởng của các chủng linh chi đột biến trên môi trường nhân giống cấp 03 chủng cho thấy khả năng sinh trưởng hệ sợi của hai chủng đột biến D-0,75, D-1,0 và D-1,25 có sự vượt trội hơn so với các chủng đột biến còn lại và giống đối chứng (Dt). Tốc độ mọc sợi của ba chủng đột biến D-0,75, D-1,0 và D-1,25 được ghi nhận là 10,01 mm/ngày, 9,8 mm/ngày và 9,6 mm/ngày, cao hơn giống đối chứng 8,8 mm/ngày. Sự sai khác này có ý nghĩa ở độ tin cậy P<0,05. Hệ sợi nấm của ba chủng đột biến D-0,75; D-1,0 và D-1,25 có màu trắng đồng nhất, phân bố đều trên cơ chất và hệ sợi trưởng thành chậm hình thành các mô sẹo.
2.3. Khả năng sinh trưởng hệ sợi, hình thành và phát triển mầm quả thể của các chủng linh chi đột biến trên giá thể nuôi trồng
Để đánh giá tiềm năng nuôi trồng, các chủng linh chi đột biến được nuôi trồng trên giá thể mùn cưa có bổ sung 10% dinh dưỡng. Tốc độ mọc sợi và đặc điểm hệ sợi của các chủng đột biến được ghi nhận ở hình 5. Hệ sợi của giống đối chứng và các chủng đột biến đều có màu trắng và phát triển đều trên bề mặt cơ chất nuôi trồng với tốc độ mọc sợi tương đối cao (hình 5). Trong các chủng đột biến khảo nghiệm, 4 chủng đột biến D-0,5, D-0,75, D-1,0 và D-1,25 cho thấy khả năng sinh trưởng hệ sợi nhanh hơn giống đối chứng (Hình 4).
Các chủng linh chi đều có khả năng hình thành và phát triển mầm quả thể trên cơ chất nuôi trồng. Thời gian hình thành mầm quả thể dạo động từ 33 đến 42 ngày và thời gian thu hoạch từ 81 đến 89 ngày. Đặc điểm hình thái quả thể (chiều dài cuống nấm và đường kính quả thể) được theo dõi. Kết quả thu được cho thấy so với giống đối chứng, các chủng đột biến có hình thái quả thể tương đối khác biệt (Hình 5,6).
Về chiều cuống nấm quả thể, các chủng đột biến có sự khác biệt lớn. Chủng D-0,25 có chiều dài cuống quả thể dài nhất, trung bình 6,7 cm, cao hơn đối chứng Dt (4,6 cm), sự khác biệt này có ỹ nghĩa ở độ tin cậy 95% (hình 5B). Thứ đến là chủng D-0,75, D-1,0, D-1,25 có chiều dài cuống quả thể trung bình, dao động từ 4,1-4,2 cm, tương đương với giống đối chứng (Dt). Các chủng D-0,5, D-1,75, D-2,0 có độ dài cuống quả thể ngắn nhất, trung bình 2,4-2,8 cm (hình 5B).
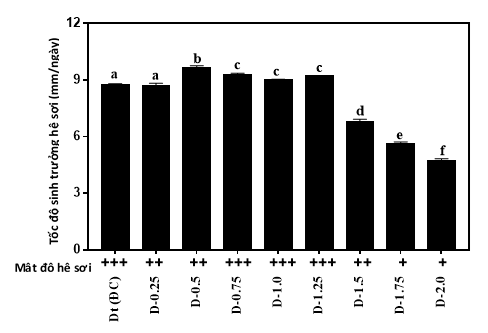 |
Hình 4. Sinh trưởng hệ sợi của các chủng linh chi đột biến trên giá thể nuôi trồng
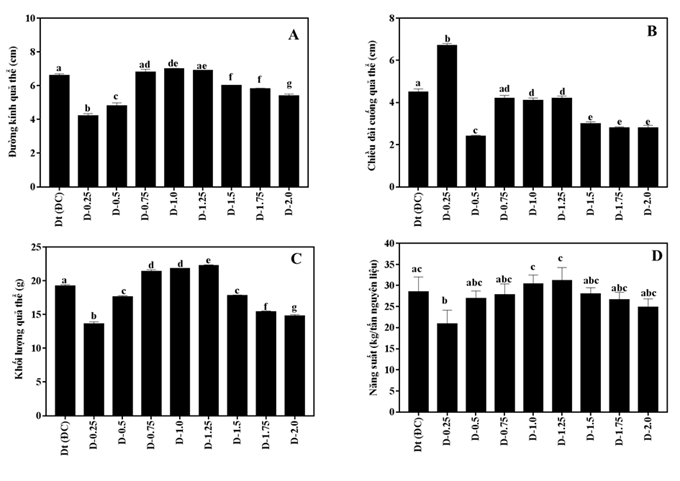 |
Hình 5. Đường kính quả thể (A), chiều dài cuống quả thể (B), khối lượng quả thể (C) và năng suất (D) của các chủng nấm linh chi đột biến và giống đối chứng
Về chỉ tiêu đường kính quả thể nấm cũng có sự biến động lớn giữa các chủng nghiên cứu. Đường kính quả thể của các chủng linh chi đột biến được chia làm 03 nhóm. Nhóm 1, gồm các chủng có đường kính quả thể lớn, đường kính trung bình đạt từ 6,6 – 7,0 cm, gồm các chủng D-0,75; D-1,0; D1,25 và giống đối chứng Dt. Đường kính quả thể của các chủng này không có sự khác biệt có ý nghĩa ở mức ý nghĩa α = 0,05. Nhóm thứ 2, là các chủng có đường kính quả thể trung bình dao động từ 5,4 – 6,0 cm, gồm các chủng D-1,50, D-1,75, D-2,0. Cuối cùng là nhóm có đường kính quả thể nhỏ nhất, trung bình chỉ đạt 4,2-4,8cm. Nhóm này gồm các chủng D-0,25 và D-0,5 (hình 5A).
Tương tự như các chỉ tiêu về đường kính quả thể và chiều dài cuống quả thể, khối lượng quả thể nấm của các chủng linh chi đột biến khảo sát cũng có sự sai khác có ý nghĩa ở độ tin cậy P ≤0,05. Trong đó ghi nhận, khối lượng quả thể của 3 chủng D-0,75, D-1,0; D-1,25 là cao nhất, trung bình đạt từ 21,4 – 22,2 g/quả, cao hơn giống đối chứng Dt (19,2 g/quả). Các chủng còn lại có khối lượng quả thể nhỏ hơn giống đối chứng, hai chủng D-0,5 và D-1,50 có khối lượng quả thể tương đương nhau (17,6 và 17,8 g/quả), các chủng D-0,25 và D-2,0 có khối lượng quả thể thuộc nhóm thấp nhất (dao động từ 13,6 – 14,8g/quả).
Năng suất nấm linh chi phụ thuộc chặt chẽ vào nhiều yếu tố ngoại cảnh: nhiệt độ, độ ẩm, oxy, ánh sáng và nồng độ CO2 [8][11]. Trong thí nghiệm này, các chủng linh chi đột biến và giống đối chứng được nuôi trồng trong cùng một điều kiện. Kết quả ở hình 6D cho thấy, các chủng linh chi đột biến được chiếu với các liều lượng bức xạ khác nhau cho năng suất nấm khác nhau. Hai chủng linh chi đột biến ở liều lượng 1,0 kGy và 1,25kGy có năng suất nấm cao nhất và khác biệt có ý nghĩa so với các chủng còn lại. Năng suất nấm linh chi khô thu được của 02 chủng D-1,0 và D-1,25 lần lượt là 30,3 kg (tăng 7,07%) và 30,5 kg (tăng 7,77%) nấm khô/tấn nguyên liệu. Chủng D-0,75 có năng suất đứng hàng thứ 3 (29,0 kg/tấn NL (tăng 2,47%), tuy nhiên lại không có sự khác biệt có ý nghĩa so với đối chứng Dt (28,3 kg/tấn NL). Các chủng D-0,5, D-1,50, D-1,75, D-2,0 có năng suất thấp hơn giống đối chứng. Chủng D-0,25 là chủng có năng suất thấp nhất (22,4 kg/tấn NL). Như vậy, trong 8 chủng linh chi đột biến, D-1,0 và D1,25 là hai chủng được đánh giá tiềm năng về năng suất và giá trị dược liệu vì vậy có thể được đưa vào khảo nghiệm nuôi trồng.
 |
Hình 6. Quả thể của các chủng linh chi đột biến D-1,0, D-1,25 và giống đối chứng Dt
3. KẾT LUẬN
Cường độ chiếu xạ có ảnh hưởng đến khả năng sinh trưởng hệ sợi và hình thái quả thể và hàm lượng dược liệu của nấm linh chi (Ganoderma lucidum). Chủng D-0,75; D-1,0; D-1,25 có tốc độ sinh trưởng hệ sợi trên môi trường nuôi cấy thuần khiết, môi trường nhân giống cấp 2 cao hơn các chủng còn lại. Trên giá thể nuôi trồng, các chủng đột biến đều cho ra quả thể. Trong đó, chủng D-1,0 và D-1,25 cho năng suất sinh học cao và lần lượt đạt 30,3 và 30,5 kg nấm khô/tấn nguyên liệu. Do đó, chủng D-1,0 và D-1,25 có thể được xem là chủng tiềm năng và đưa vào khảo nghiệm nuôi trồng.
NCS. Trần Thu Hà, khoa Công nghệ sinh học
TÀI LIỆU THAM KHẢO
1. Beaulieu. M., Daprano. M.B.G. and Lacroix. M. (1999). Dose rate effect of γ irradiation on phenolic compounds, polyphenol oxidase, and browning of mushrooms (Agaricus bisporus). Journal of Agricultural and Food Chemistry. 47: 2537-2543.
2. Djajanegara, I. R. A. (2008). White oyster mushroom (Pleurotus florida) mutant with altered antioxidant contents. Biotropha. 15(1): 65–72.
3. Lee, Y. K., Chang, H. H., Kim, J. S., Kim, J. K. and Lee, K. S. (2000). Lignocellulolytic mutants of Pleurotus ostreatus induced by gamma-ray radiation and their genetic similarities. Radiation Physics and Chemistry, 57(2), 145–150.
4. Nakagawa, T., Zhu, Q., Tamrakar, S., Amen, Y., Mori, Y., Suhara, H., Kaneko, S., Kawashima, H., Okuzono, K., Inoue, Y. and Ohnuki K. (2018). Changes in content of triterpenoids and polysaccharides in Ganoderma lingzhi at different growth stages. Journal of Natural Medicines. 72(3): 734-744.
5. Nguyen, B.T, Ngo, X. N., Le, V.V., Nguyen, T.L., Ry. K., Nguyen, D.H (2019). Optimal culture conditions for mycelial growth and fruiting body formation of Ling Zhi mushroom Ganoderma lucidum strain GA3. Vietnam Journal of Science, Technology and Engineering. 61(1): 62-67.
6. Nguyễn Hữu Đống, Đinh Xuân Linh, Nguyễn Thị Sơn, Ngô Xuân Nghiễn và Zani Federico (2005). Nấm ăn – Cơ sở khoa học và công nghệ nuôi trồng, Nhà xuất bản Nông nghiệp, Hà Nội.
7. Sathesh-prabu, C., & Lee, Y. (2011). Breeding and Genetic Improvement of Mushrooms. Radiation Oncology Journal. 5(4): 285–295.
8. Skalicka–Woźniak, K., Szypowski, J., Łoś, R., Siwulski, M., Sobieralski, K., Głowniak, K. and Malm, A. (2012). Evaluation of polysaccharides content in fruit bodies and their antimicrobal activity of four Ganoderma lucidum (W. Curt.: Fr.) P. Karst. Strains cultivated on different wood type substrates. Acta Societatis Botanicorum Poloniae. 81: 17-21.
9. Stamets, P. (1993). Ganoderma lucidum (Wm, Curtis: Fries) Karsten. in growing gourmet and medicinal mushrooms. In: Stamets P. (Ed.). Ten Speed Press, Berkeley, CA. 355-369.
10. Trịnh Tam Kiệt (2012). Nấm lớn ở Việt Nam. Tập 2. Nhà xuất bản Khoa học Tự nhiên và Công nghệ, Hà Nội.
11. Zhou, X. W., Su, K. Q. and Zhang, Y. M. (2012). Applied modern biotechnology for cultivation of Ganoderma and development of their products., Applied Microbiology and Biotechnology. 93(3): 941-963.